SISTEMATICA
INTRODUZIONE
Un leggero chiarimento sulla comparsa delle mandibole e mascelle dal punto di vista temporale (2023. Per la fonte, clicca qui)
Mascelle e mandibole: 3 diverse teorie a confronto
Gli gnatostomi sono un gruppo di pesci molto diversificato dai parenti agnati, ovvero dai pesci privi di mandibole e mascelle. Tra i tanti caratteri, questa è quella che salta subito all'occhio in quanto parliamo di una struttura complessa che sembra comparire all'improvviso nel record fossile, magari per via di geni hox, Evo-Devo, e/o di macromutazioni e chi più ne ha più ne metta. Al momento, l'unica cosa che sappiamo è che un grande cambiamento ha coinvolto molte regioni anatomiche che si sono trasformate in una struttura capace di "aprirsi" e di "chiudersi", e che in tempi successivi verrà ereditata da ogni vertebrato. Quello che sappiamo è che gli gnatostomi sono diretti discendenti degli agnati. Vediamo quali sono gli studi in merito.
Teoria classica (Gegenbaur, 1872)
Le mandibole e le mascelle si sarebbero originate da una delle arcate branchiali anteriori, quelle più vicine al cranio, ma non ci sono 'fossili' in merito, nemmeno di specie con caratteristiche intermedie. Possiamo riassumere il processo così:
1)Il primo arco branchiale avrebbe dato origine a parte del Neurocranio;
2)Il secondo arco avrebbe dato origine al Palatoquadrato, ovvero alla parte principale della mascella, e alla cartilagine di Meckel (il nucleo delle mandibole);
3)Il terzo avrebbe formato elementi di giunzione tra mascelle e mandibole, conosciuti come Iomandibolare e Ceratoide.
La mancanza di fossili crea qualche problema in quanto sono presenti morfologie complesse e derivate negli gnatostomi, ma mancano fossili "intermedi" quindi è difficile capire quali strutture si sono modificate in altre, ma in linea di massima con la "teoria classica" si concorda che l'osso ioide e le mascelle derivino da archi branchiali modificati, così come la Tromba di Eustachio per i vertebrati terrestri ,o spiracolo per i pesci, potrebbe essere stato un solco faringeo (o fessura branchiale)
Teoria composita
L'arcata mandibolare si sarebbe formata da elementi di più arcate branchiali, adiacenti, che avrebbero contribuito anche alla formazione del neurocranio. In questo caso, il palatoquadrato e la cartilagine di Meckel potrebbero non essere comparsi con la modificazione delle arcate branchiali, ma potrebbero essere comparsi precedentemente già negli agnati (ed associati già alla bocca).
Per quanto riguarda queste due teorie, gli archi branchiali degli agnati sono posti esternamente, e non sono omologhi agli archi branchiali degli gnatostomi (che sono posti internamente). Devono essere avvenute delle modificazioni dopo il "passaggio" da archi esterni ed interni.
Ora, però, la storia si fa più interessante in quanto ci sono diversi studi che hanno provato a spiegare queste modificazioni appena citate.
Mallatt (1984) ipotizza "una nuova bocca"
Questo studio si basa sulla 'non-omologia' degli archi branchiali tra agnati (le lamprede possiedono archi posizionati lateralmente) e gli gnatostomi (con gli archi posizionati medialmente).
Mallatt afferma comunque che dagli archi branchiali si sarebbero originate le mascelle e le mandibole, senza spingersi oltre alla complessa modificazione dei diversi archi branchiali visti nella "teoria classica". Infatti, afferma che la modificazione degli archi sarebbe legata ad una ventilazione più efficiente, in quanto l'arco anteriore (il primo), ingrossandosi, risulterebbe più mobile e quindi potrebbe aver svolto una funzione simile, o molto vicina, a quella della bocca come la intendiamo noi.
Infatti, lui nota che gli agnati fossili, come anche le lamprede, possiedono quelle che volgarmente possiamo definire labbra, e guance, che racchiudono una cavità orale. Possiamo riassumere quest'ipotesi così:
-Davanti la faringe, quindi dentro o dietro la bocca, si sarebbero formate le mascelle e le mandibole;
-La posizione risulterebbe essere vantaggiosa perché si possono creare "vortici d'acqua" atti alla predazione (in questo modo si risucchiano le prede, e non è diverso placodermi, uno dei primi gruppi di gnatostomi, in quanto erano capaci di creare vortici d'acqua chiudendo e aprendo la bocca);
-Grazie a meccanismi evolutivi, come la Selezione Naturale (ma non è detto, non avendo molte prove fossili a riguardo), questo carattere potrebbe essere stato ereditato da più gruppi, subendo mutazioni/modificazioni come la riduzione della bocca tra le mascelle/mandibole e le labbra.
Ecco, in questo modo abbiamo la "nuova bocca" di Mallatt, una struttura che non sarebbe comparsa con gli gnatostomi ma che si sarebbe sviluppata da una "bocca precedente".
In conclusione, possiamo aggiungere alcuni risultati interessanti (anche se non sono supportati da fossili con caratteristiche intermedie):
-L'ulteriore specializzazione degli archi (che si sono modificati in mascelle), deriva dalla nutrizione tramite suzione;
-La faringe potrebbe essere un carattere exaptato (ci si riferisce all' 'exaptation', un meccanismo evolutivo conosciuto anche volgarmente come "preadattamento". Sostanzialmente, si tratta di un carattere che è codificato geneticamente per svolgere determinate funzioni, come le penne e le piume per quanto riguarda la termoregolazione, ma può anche svolgere altre funzioni non legate alla genetica. Per esempio, penne e piume svolgono un ruolo importante per il volo). Sostanzialmente, la faringe permetteva di nutrirsi tramite 'suzione', mentre in organismi più derivati svolge un ruolo nella respirazione.
Janvier (1996) e lo 'scheletro velare'
Questo è uno studio già più "recente" rispetto a quello di Mallatt, e ci si basa un po' di più su evidenze biologiche e non fossile. Infatti, evidenze genomiche indicano che l'arco mandibolare è omologo al cosiddetto "velo" delle lamprede. In parole povere, con velo si intende una membrana che rende separato il cavo orale all'interno della faringe. Come se fosse una sezione a parte. Analizziamo punto per punto questo studio:
-Nelle lamprede, il velo è sostenuto dalle cartilagini velari;
-Janvier sostiene che gli archi mandibolari siano omologhi a queste cartilagini;
-Le cartilagini velari possiedono porzioni ventrali e dorsali.
In conclusione, per quanto riguarda le teorie appena viste, possiamo dire che Mallatt ipotizza che i vertebrati ancestrali avessero archi branchiali sia interni che esterni, soltanto che negli gnatostomi si sono persi quelli esterni, mentre in agnati più derivati (come le lamprede) si sono persi quelli interni.
Per Janvier, invece, gli archi interni degli gnatostomi sono il risultato di alcune trasformazioni degli elementi mediali dello scheletro velare. (da mediali a interni).
Chi ha ragione tra Mallatt e Janvier?
Questa volta ad intromettersi sono Sleight & Gills (2020), in quanto hanno compiuto studi ontogenetici che riguardano le branchie di una razza. Prima di poter andare avanti, dobbiamo chiarire un paio di punti:
-Le branchie degli gnatostomi e delle lamprede derivano dall'endoderma (circondato a sua volta da ectoderma e cresta neurale);
-Negli gnatostomi, gli archi branchiali mediali vengono prodotti dalla cresta neurale, ma il processo viene "innescato" dall'endoderma del tubo digerente;
-Nelle lamprede, invece, il fattore scatenante è l'ectoderma, e di conseguenza si avrà una posizione diversa degli archi branchiali.
Ritornando allo studio di Sleight & Gills, è stato dimostrato che in una razza gli archi neurali sono composti posteriormente dal mesenchima (in generale, viene dato per assodato che dal mesenchima mesodermico derivino le 'pinne pari'. Quelle, per esempio, del "cinto pettorale" dei pesci, o comunque quelle pinne che si sviluppano in coppia), e anteriormente dalla cresta neurale
Sostanzialmente, questo studio sembra dare ragione a Janvier, mentre Mallatt parrebbe essere supportato da "Metaspriggina walcotti" e dai suoi, probabili, archi branchiali. Questo fossile è stato rinvenuto nel famosissimo lagerstätte conosciuto come Burgess Shale (se non avete letto "La vita Meravigliosa di Gould, dovete farlo al più presto. Riassumiamo qualche caratteristica:
-Presenta un corpo compresso lateralmente e fusiforme;
-Il corpo si restringe man mano che ci si avvicina al 'capo', che non è separato dal resto del corpo;
-Si notano piccoli occhi e un'apertura buccale;
-Presenta una notocorda relativamente cilindrica (forse a causa di fenomeni tafonomici), che si estende dal capo alla punta della coda;
-Dal punto di vista filogenetico, appartiene al Phylum Chordata e non è troppo distante dai vertebrati.
La preservazione eccezionale ci permette anche di studiare tratti anatomico-organici che difficilmente si preservano (tranne, appunto, nei Lagerstätte). In questo caso sembrano esserci narici esterne pari (un carattere distintivo degli gnatostomi, o comunque in generale dei vertebrati dotati di mascelle/mandibole). Nella regione centrale del corpo, invece, sono state trovate tracce di 'barre cartilaginee' che potrebbero corrispondere, probabilmente, agli archi branchiali.
Naturalmente, su questo fantastico ritrovamento ci sarebbe molto da dire, ma gli ultimi elementi bastano per dare, forse, ragione anche a Mallatt.
Detto questo, voi cosa ne pensate? Qual è la teoria (scientifica) che vi affascina di più sull'evoluzione delle mandibole e delle mascelle?
ACANTHODII
PLACODERMI
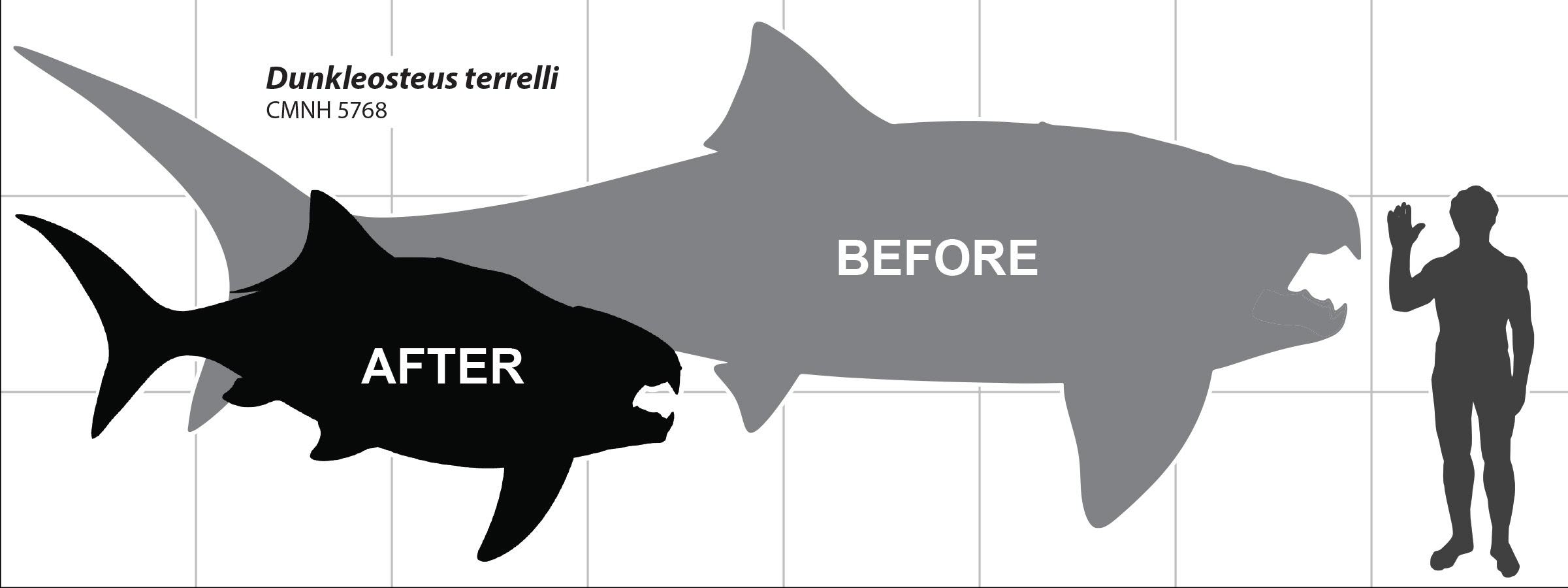
La sovrastima della taglia della specie Dunkleosteus terrelli (per la fonte, clicca qui)
La specie Dunkleosteus terrelli è un placoderma molto antico, conosciuto sia per le sue peculiarità morfologiche che per la sua grande 'stazza'. Infatti, in ogni rappresentazione dei mari del Devoniano (416-359,5 milioni di anni fa circa), troviamo questo bestione che attacca come un forsennato a destra e a manca qualsiasi tipo di preda, riconosciuto anche come primo (o uno dei primi) predatore vertebrato all'apice della catena alimentare.
Ora ritorniamo al nostro D. terrelli. Ma è stato davvero un predatore gigantesco? Questa ricerca del 2023 indica che gli studi precedenti sovrastimarono questa specie, che risulta essere molto più ridotta del previsto, anche se ancora non si conoscono le reali dimensioni in quanto l'armatura toracica e la testa sono, al momento, gli unici elementi ossei ad essere presenti nel registro fossilifero.
Le "vecchie" stime indicavano una lunghezza di questo pesce di circa 5-10 m, che sono perlopiù qualitative e non derivanti da analisi statistiche, a differenza di questo studio. Infatti, è stata utilizzata come parametro la lunghezza orbita-opercolare (o meglio, per alcuni pesci fino a dove finiscono le fessure branchiali in quanto quest'osso è assente in alcune specie analizzate), comparato con quello di altri artrodiri e di pesci odierni.
Il risultato è che la lunghezza orbita-opercolare è fortemente correlata con la lunghezza totale dei pesci, e ciò permette di predire con precisione la lunghezza di altri artrodiri (ordine Arthrodira, a cui appartiene questa specie). Per il nostro amico D. terrelli si ottengono, come detto prima, dimensioni un po' più contenute rispetto ai 5-10 m calcolati in precedenza:
-Circa 3,4 m è/era la lunghezza di un adulto nella norma;
-Per gli individui più grandi conosciuti si arriva a circa 4,1 m.
In conclusione, si può dire che i grandi artrodiri (Dunkleosteus, Titanichthys) erano molto più piccoli di quanto si pensasse, non raggiungono superiori superiori ai 5 m (fino al Carbonifero), e che la forma del corpo (oltre ad essere corta) è anche più cilindrica, se vogliamo un po' più "tozza", e ciò li rende esteriormente diversi dagli elasmobranchi o dagli attinopterigi (che presentano in genere un corpo un po' più affusolato).
Paleobiologia dei placodermi in linea generale
Il primo cuore fossile tridimensionale e la prima traccia di guaina mielinica nel record fossile (per la fonte, clicca qui, qui e qui)
Una notizia che ha fatto scalpore è il ritrovamento di un cuore fossile datato circa 380 milioni di anni fa. Ma, la cosa sorprendente, è che sono stati rinvenuti anche il fegato, l'intestino e lo stomaco.
Già la fossilizzazione di "parti dure" è un evento raro, ma rinvenire organi fossili è ancora più raro, e se riusciamo a studiarli è grazie proprio ai lagerstätte, un particolare corpo sedimentario che in particolari condizioni permette alla materia organica "molle", che in genere si degrada in poco tempo, di preservarsi (sotto forma di roccia, naturalmente!).
Ecco quali informazioni anatomiche ci fornisce il fossile di placoderma arthrodira rinvenuto recentemente:
- E' al momento l'unico cuore fossilizzato tridimensionalmente. La struttura si presenta schiacciata e a forma di 'S', ed è ben separato dal fegato e da altri organi addominali;
-Lo stomaco possedeva delle pareti spesse;
Sostanzialmente, la disposizione e la forma del cuore indicano che la presenza di un "collo", o comunque una proto-struttura che permettesse di movimento del capo, a livello filogenetico comparve ben prima di quanto studiato precedentemente.
Fonte:
science.orgUn altro ritrovamento paleobiologico molto importante, forse passato in sordina qualche tempo fa, è la scoperta di tracce di guaina mielinica, la prima nel record fossile ed associata ai placodermi. La maggior parte dei vertebrati è caratterizzata dalla presenza di guaina mielinica che consente grandi aumenti della velocità di propagazione dell'impulso lungo gli assoni, ed il gruppo candidato è quello dei placodermi per diversi motivi:
-Sono i primi pesci a presentare una morfologia comune a tutti i vertebrati (tranne missine e lamprede), cioè le mandibole e le mascelle. In pratica, furono i primi vertebrati a riuscire a chiudere e aprire la bocca grazie ad un'articolazione a "cerniera", posta tra lo scudo toracico e lo scudo cefalico che permetteva a quest'ultimo, quindi alle mandibole e mascelle, di aprirsi e chiudersi per catturare le prede. Una struttura abbastanza complessa che solo grazie alla presenza di guaina mielinica poteva essere "controllata" in modo efficiente;
-Sono stati scoperti fossili di placodermi conservati in modo eccezionale, in cui è possibile vedere degli elementi che confermerebbero la presenza della guaina mielinica, e che di conseguenza indicherebbero una probabile comparsa della stessa nel Devoniano.
Nell'immagine con quella sorta di "struttura gialla", si possono notare le impronte dei nervi cranici e dei forami che attraversano i crani di questi pesci. I diametri dei nervi oculomotori sono molto simili a quelli degli osteostraci (pesci senza mascelle del Paleozoico estinti), mentre la lunghezza dei nervi era 10 volte maggiore indicando che, pur mantenendo un diametro costante, il sistema oculomotore deve essere stato mielinizzato per "funzionare" come una via motoria a conduzione rapida, cioè gli assoni dovevano essere già avvolti da guaina mielinica per far "correre" piu velocemente l'impulso/informazione.
La seconda immagine, quella in bianco e nero con le frecce, rappresenta un 'neurone fossile' con piastre terminali motorie (indicate dalle frecce) che si attaccano alle fibre muscolari. È una prova indiretta di neuroni con guaine mieliniche.
I
mpronte dei nervi cranici e dei forami. Fonte immagine science.org
Assone fossile. Fonte:
science.org




















.jpg)

.jpg)


